
THE NATURAL ABUNDANCE OF L-CANAVANINE, AN ACTIVE ANTICANCER AGENT, IN ALFALFA, MEDICAGO SATIVA (L.).
GERALD A.ROSENTHAL AND PALESA NKOMO
Laboratory of Biochemical Ecology, University of Kentucky, Lexington, KY 40506
----------------------------------------------------------------------------------------------------------------------------
SUMMARY--L-Canavanine, a potentially toxic antimetabolite of L-arginine stored by many leguminous plants, has demonstrative antineoplastic activity against a number of animal-bearing carcinomas and cancer cell lines. This investigation evaluated the natural abundance of this anti-cancer compound in commercially-available sprouts, and in ten varieties of the seed of alfalfa, Medicago sativa (L.). While the abundance of canavanine (on a dry matter basis) in the commercially-grown sprouts varied according to the source, this plant stored appreciable canavanine with a content that ranged from 1.3 to 2.4% of the dry matter. The seeds were also rich in this nonprotein amino acid and ranged from 1.4 to 1.8% of the dry matter, and had a mean + SEM canavanine content value of 1.54 + 0.03%. No evidence was found for elevated canavanine content as a result of seed germination and early growth. The canavanine content of the alfalfa seeds was comparable to the levels found in the seeds of several members of the genus Canavalia, which are amongst the more abundance sources of this antimetabolite. -----------------------------------------------------------------------------------------------------------------------------
INTRODUCTION
The nonprotein amino acid, L-canavanine is a potent arginine antimetabolite stored by many leguminous plants (Bell, 1958; Bell et al., 1978) as part of their chemical barrier against predation and disease-causing organisms (Rosenthal, 1977a; Rosenthal, 1992).
This arginine antagonist also elicited antineoplastic effects against a number of carcinomas. This observation was made initially by Kruse and McCoy (1958) who reported that canavanine competed with arginine in meeting the growth requirements of Walker carcinosarcoma 256 cells. Kruse et al. (1959) subsequently demonstrated that canavanine was incorporated into the proteins of these cancer cells, and that the diminution in the amount of arginine in the protein hydrolysate equaled the canavanine content.
A detailed study of canavanine's anticancer activity was conducted with mice bearing L1210 leukemic cells (Green et al., 1980). These workers reported that DNA synthesis fell to only 9% of the control level, as assessed by [3H]thymidine incorporation, after 12-hourly i.p. injections of 20 mg canavanine per injection. In a follow-up study, Green and Ward (1983) reported that canavanine enhanced significantly the efficacy of gamma-irradiation of cultured HT-29 cells, a human tumor cell line. The lethal effect of this radiation was augmented both when canavanine was provided prior to as well as after gamma-irradiation. These workers provided convincing experimental evidence for their contention that canavanine's lethal effect was manifested preferentially in rapidly proliferating cells--a property essential to chemotherapeutic efficacy (Green and Ward, 1983).
Canavanine disrupted the growth of a colonic carcinoma in male Fischer rats (Thomas et al., 1986). Providing 3.0 g kg-1 canavanine by parenteral injection to animals bearing palpable tumors, resulted in a 13% reduction in tumor volume after 5 treatment days (Thompson et al., 1986). Because of the preferential uptake of L-[guanidinooxy-14C]canavanine into the proteins of the pancreas (Thomas and Rosenthal, 1987), canavanine's ability to affect the growth of MIA PaCa-2, a human pancreatic adenocarcinoma cell line, was assessed. Exposing these cells to canavanine, in a minimal arginine-containing media, resulted in significant cell death and an IC50 value of 10 mM for canavanine (Swaffar et al., 1994).
Given the growing evidence of the anti-cancer efficacy of canavanine, we re-opened the question of how effectively alfalfa, Medicago saliva (L.), provided as commercially-available sprouts, served as a dietary source of canavanine . We also assessed the variation in seed canavanine content by carefully determining its natural abundance in 10 varieties.
Prior study of canavanine abundance in this plant was limited to the simple observation of Bell (1958) that alfalfa seeds available in the United Kingdom contained 1.46% canavanine by dry weight. No data were provided on how this value was determined nor were common articles of everyday diet evaluated.
METHODS AND MATERIALS
Materials. Fresh alfalfa sprouts were provided by a local Kroger, Inc. supermarket on three occasions From late June through mid-July of 1996. Multiple fresh sprout samples were also obtained at this time from New Natives, a grower in Aptos, CA, and the Good Food Cooperative of Lexington, KY. Sprouts were also grown from seed in the greenhouse of the University of Kentucky under ambient conditions in July. Seeds of the following varieties of M. sativa were obtained from Sphar & Co. (a division of Scott Seed Co.), Winchester, KY: Pioneer Hi-Bred 5454L and 5373L, ABT 405, ABT Supercuts, Aggressor, Alfagraze, Scott Brand ProBlend, Affinity + Z. Amerigraze, and Total + Z. Pioneer Hi-Bred International, Inc., Johnston, Iowa provided ten seed varieties (see Table 2). Chicago's Indoor Gardens provided two seed varieties.
Chemicals and Biochemicals. L-Canavanine was isolated from jack bean seeds, Canavalia ensiformis, and purified by re-crystallization as described elsewhere (Bass et al., 1995). Automated amino acid analysis established the absence of contaminant amino acids in the canavanine. NMR, elemental analysis, melting point, and optical rotation values were within established parameters (Bass et al., 1995). All reagents were obtained from Fisher/Agros Corp. or Sigma/Aldrich Chemical Co.
Preparation of the Plant Extract. Twenty g of alfalfa sprouts, either fresh or frozen at -20oC, were ground with a Servall Omni-mixer at full power for 3 min with 60% aqueous ethanol containing 1%(v/v) HCI. The slurry was decanted into a centrifuge bottle along with two washes of the mixing chamber. After centrifugation at 12,200 xg for 25 min. the supernatant solution was decanted into a volumetric cylinder. This extraction procedure removed 95 + 1% of the canavanine of the plant sample as determined by stirring the pellet in the above solvent overnight at 3°C.
Canavanine Verification. About 20 mL of the final extract was taken to pH 7.6 with N NaOH and filtered over Whatman no. 1 paper. The filtrate was concentrated by rotary evaporation in vacuo, and the residue suspended in deionized water and evaporated as above. The residue was taken up in 10 mL of 50 mM tricine buffer (pH 7.6) containing 1 mM MnCI2 and 0.1% (v/v) mercaptoethanol. After filtering the suspension as above, 0.4 mL of the filtrate was reacted overnight at 37°C with 100 mL of purified arginase, prepared as described elsewhere (Rosenthal, 1977b). The purified arginase sample contained sufficient enzyme to degrade a 5-fold excess of the amount of canavanine thought to be in the sample. Deionized water (0.5 mL) was added to the reaction mixture which was then evaluated colorimetrically for canavanine.
These assessments, insuring that canavanine was responsible for the magenta, PCAF-chromogen, always resulted in a complete loss of the PCAF-canavanine chromogen.
A 25 mL sample of the plant extract was dispensed into a centrifuge tube and treated with 0.85 mL of 4N NaOH. After mixing thoroughly, the turbid solution was clarified by centrifugation at 27,000 xg for 15 min. Colorimetric analysis, in triplicate, was conducted on an appropriate sample of the final supernatant solution (Rosenthal, 1977b). All reported values are the mean + SEM of 4 separate determinations for the sprouts and 3 determinations for the seeds.
Colorimetric analysis. The plant extract (1.0 mL) was treated with an equal vol of 200 mM potassium phosphate buffer (pH 7.0) and 0.2 mL of 1.0% (w/v) potassium persulfate. After vortexing vigorously, 100 mL of 1.0% (w/v) pentacyanoammonioferrate (PCAF) solution was added, the tube agitated vigorously once again, and color development allowed to proceed for at least 30 min. The PCAF-canavanine chromogen was read at 530 nm where chromogen formation obeys Lambert-Beer's law to 0.8 mM. PCAF was prepared from recrystallized commercial sodium nitroprusside dihydrate after the method of Fearon (Fearon, 1946).
Automated Amino Acid Analysis. Automated amino acid analysis was conducted with a Dionex D-300 Automated Amino Acid Analyzer utilizing commercial lithium citrate (Pickering) gradient elusion and ninhydrin detection at 570 nm. These analyses were not intended to quantitate the amount of a particular amino acid in a given sample, but rather to determine to abundance of canavanine relative to the other amino acids present in the sample.
Dry Weight Determination. The sprout or seed dry weight for M. sativa was determined by drying at 60°C in a convection drying oven (Precision Scientific, Inc., Model 18) for 2 d. The Canavalia seeds were pulverized mechanically prior to dry weight evaluation. Recovery The recovery of canavanine by the standard extraction procedure was established by the addition of 5 mCi of L-[guanidinooxy-14Clcanavanine to the plant material. Evaluation by liquid scintillation spectroscopy revealed that 97% of the radiolabeled canavanine was recovered in the final extract taken for calorimetric analysis.
RESULTS AND DISCUSSION
Sprouts.
Analysis of alfalfa sprouts revealed that while the concentration of canavanine varied in this table legume, it was an excellent source of canavanine. (Table 1). The sprouts obtained directly from the commercial grower contained on average 1.3% canavanine by dry weight. These samples of alfalfa sprouts were heavily laden with seed coats, and they possessed on average 6.7% dry matter. The sprouts obtained from the local supermarket contained 1.4% canavanine by dry weight. These samples also contained seeds coats, and the sprouts had an average dry weight of 9.7% (Table 1).
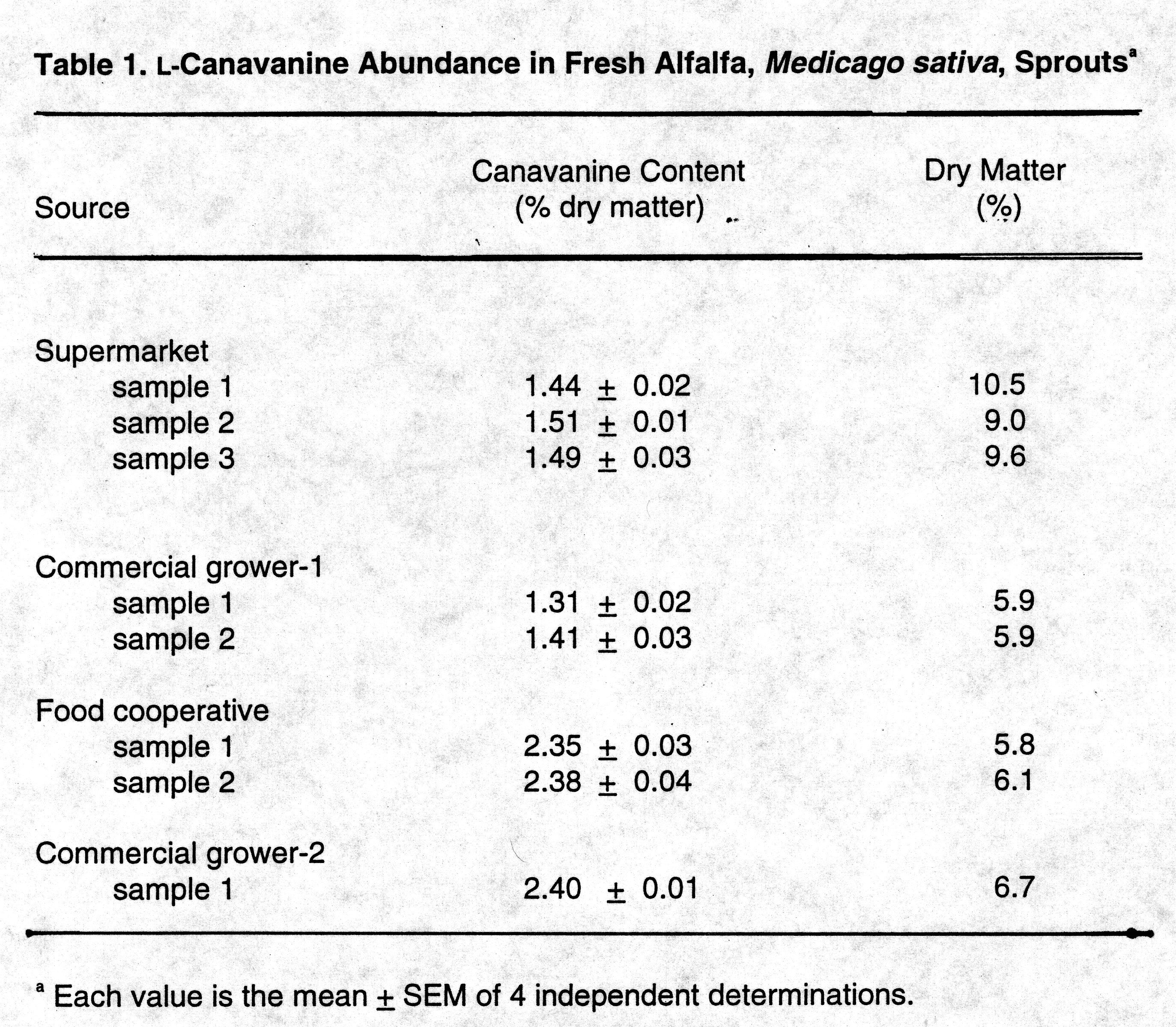
On a fresh weight basis, this represents a significant variation in canavanine abundance as the latter source of alfalfa contained more than half again as much canavanine as the former. The final samples of sprouts, purchased from the food cooperative, was washed free of all testes. These samples contained 2.5% canavanine by dry weight, and had an average 5.95% dry matter. Removal of the spent testa in the food cooperative material increased the proportion of fresh sprout in the sample, and decreased the dry matter content. These factors elevated the canavanine abundance, and contributed to the overall canavanine richness of their samples.
To establish if the canavanine richness of the sprouts also reflected a high natural abundance in the seeds used to prepare these sprouts, the source of this commercial product was traced to Chicago's Indoor Garden. Analysis of the appropriate seeds revealed a canavanine content of 1.43 + 0.02% of the dry matter which is not greater than that found in the alfalfa seed samples tested in this study (see below).
Seeds.
The canavanine content of the alfalfa seed was predicated upon an evaluation of 10 varieties of seed provided by Pioneer Hi-bred (Table 2). This grower developed their varieties for resistance to insect infestation, hardiness, high yield, and overall robustness. In a preliminary screening of 10 varieties provided by Sphar Seed Co., the two Pioneer Hi-bred varieties were the richest in canavanine of the 10 varieties initially evaluated. Subsequent analysis of 10 varieties of the Pioneer Hl-Bred seeds revealed a canavanine content on a dry weight basis, whose mean + SEM equaled 1.54 + 0.03%, and ranged from 1.4 to 1.8% (Table 2).

Canavanine in Canavalia. In order to more realistically evaluate the natural abundance of canavanine in M. saliva, the canavanine content of the seeds was compared to a number of species of Canavalia, which are rich sources of canavanine (Figure 1). This comparative evaluation disclosed that while M. sativum contained less seed canavanine than the studied members of Canavalia, its natural abundance is not trivial.

Fig. 1. The relative abundance of seed canavanine in M. sativa and several members of Canavalia. The seeds were processed as described in the text. Each value is the mean + SEM for 3 independent determinations.
ACKNOWLEDGMENTS
We gratefully acknowledge the support of the Nationa! Science Foundation (Grant IBN-9302875) and an NSF REU grant to P. Nkomo. Materials required for this study were provided kindly by Krogers Supermarkets, Inc., Sphar Seed Co., Pioneer Hi-Breed Seed Co., New Natives, and Chicago's Indoor Gardens.
REFERENCES
1. Bass, M.; Crooks, P. A.; Levi, H.; NaPhoket, S.; Trifonov, L. S. and Rosenthal, G. A.(1995) Large scale production and chemical characterization of the protective, higher plant allelo-chemicals: L-canavanine and L-canaline. Biochem. Syst. Ecol. 23: 717-721.
2. Bell, E. A. (1958) Canavanine and related compounds in the Leguminosae. J. Biochem. 70, 617-619.
3. Bell, E. A.; Lackey, J. A. and Polhill, R. M. (1978) Systematic significance of canavanine in the Papilionoideae (Faboideae). Biochem. Syst. Ecol. 6: 201-212.
4. Fearon, W. R. (1946) The amidine-pentacyanoferrate reaction: a test for guanidines, urea and thio-ureas. Analyst 71: 562-566.
5. Green, M. H.; Brooks, T. L.; Mendelsohn, J. and Howell, S. B. (1980) Antitumor activity of L-canavanine against L1210 murine leukemia. Cancer Res. 40: 535-537.
6. Green, M. H. and Ward, J. F. (1983) Enhancement of human tumor cell killing by L-canavanine in combination with g-irradiation. Cancer Res. 43: 4180-4182.
7. Kruse, P. F., Jr. and McCoy, T. A. (1958) The competitive effect of canavanine on utilization of arginine in the growth of Walker carcinosarcoma 256 cells in vitro. Cancer Res. 18: 279-282.
8. Kruse, P. F., Jr.; White, P. B.; Carter, H. A. and McCoy, T. A. (1959) Incorporation of canavanine into protein of Walker carcinosarcoma 256 cells cultured in vitro. Cancer Res. 19: 122-125.
9. Miersch, J. 1992. Possible function of canavanine during germination of alfalfa seeds Medicago sativa (L.) in Guanidino Compounds in Biology and Medicine, (P. P. De Deyn, B. Marescau, V. Stalon, and A. Qureshi, eds.) John Libbey & Co. London, UK.
10. Nakatu, S.; Haratake, S.; Sakuri, Z.; Zyo, N.; Nishihara, Z. and Hayasida, M. (1964). The change of quantity of canavanine in leguminous plants in the process of germination, growth, and fruitification. Seikagaku (J. Jap. Biochem. Soc.) 36: 467-471.
11. Rosenthal, G.A. (1970) Investigations of canavanine biochemistry in the jack bean plant, Canavalia ensiformis (L.) DC. Plant Physiol. 46: 273-276.
12. Rosenthal, G. A. (1977a) The biological effects and mode of action of L-canavanine, a structural analogue of L-arginine. Q. Rev. Biol. 52: 155-178.
13. Rosenthal, G. A. (1977b) Preparation and colorimetric analysis of L-canavanine. Anal. Biochem. 77: 147-151.
14. Rosenthal, G.A. (1977c) Nitrogen allocation for L-canavanine and its relationship to chemical defense of the seed. Biochem. System. Ecol. 5: 219-220.
15. Rosenthal, G.A. (1990) Metabolism of L-canavanine and L-canaline in leguminous plants. Plant Physiol. 94: 1-3.
16. Rosenthal, G. A. (1992) L-Canavanine and chemical defense in higher plants. in Frontiers and New Horizons in Amino Acid Research; (K. Takai, ed.) Elsevier: New York.
17. Rosenthal, G.A.; Berge, M.; Ozinskas, A. and Hughes, C.H. (1988) Ability of L-canavanine to support the nitrogen metabolism in the jack bean, Canavalia ensiformis (L.) DC. J. Food Agr. Chem. 36: 1159-1163.
18. Swaffar, D. S.; Ang, C. Y.; Desai, P. B. and Rosenthal, G. A. (1994) Inhibition of the growth of human pancreatic cancer cells by the arginine antimetabolite, L-canavanine. Cancer Res. 54: 6045-6048.
19.Thomas, D. A. and Rosenthal, G. A. (1987) Metabolism of L-[guanidinooxy-14C]-canavanine in the rat. Toxicology & Appl. Pharm. 91: 406-414.
20. Thomas, D. A.; Rosenthal, G. A.; Gold, D. V. and Dickey, K. (1986) Growth inhibition of a rat colon tumor by L-canavanine. Cancer Res. 46: 2898-2903.
.